

◆Tadashi Isa, the 27th Recipient of the Tokizane Award in 2025
◆Announcement of the 2025 JNS-CNS Travel Award Program Awardees
◆[Publication] Neuroscience News FY2025 No.1
◆Awardee of 2025 Joseph Altman Award in Developmental Neuroscience, Tomohisa Toda
◆ Announcement of the 27th Recipient of the Tokizane Award in 2025
◆Announcement of the Awardees of the 2025 Neuroscience Research (NSR) Paper Awards.
◆Awardee of the 2025 Joseph Altman Award in Developmental Neuroscience
◆Promotional video for Neuroscience Research is now available on our website.
◆The Winners of the Brain Prize 2025
◆THE BRAIN PRIZE 2025 WINNERS ANNOUNCED ON MARCH 5TH (CET)
◆[Publication] Neuroscience News FY 2024 No.4
◆Participating Report of the 17th Annual Meeting of Chinese Neuroscience Society (CNS 2024)
◆“Meet the Editors” Session (July 24, 25 and 26, 2024) will be held at NEURO2024
◆Neuroscience Research Special Issue(Vol.197) is available for free now
◆Neuroscience Research is now Full Gold Open Access journal
◆Message from the President
◆Atsushi Nambu, The 26th Recipient of the Tokizane Award in 2024
◆Awardee of 2024 Joseph Altman Award in Developmental Neuroscience, Shawn Sorrells
◆Announcement of the 26th Recipients of the Tokizane Prize in 2024
◆Announcement of FENS Forum 2024 Travel Award Awardees
◆Announcement of the 2024 JNS-CNS Travel Award Program Awardees
◆Awardee of the 2024 Joseph Altman Award in Developmental Neuroscience
◆The Winners of the Brain Prize 2024
◆THE BRAIN PRIZE 2024 WINNERS ANNOUNCED ON MARCH 5TH (CET)/ 6TH MARCH(JST)
◆FENS Summer School 2023 Report (Owusu Mensah Richard Nana Abankwah)
◆Call for applications: Chief Editors of Neuroscience
◆Message from the President 'Our Neuroscience Society'
◆Awardee of 2023 Joseph Altman Award in Developmental Neuroscience, Tomasz Nowakowski
◆Announcement of the 24th Recipients of the Tokizane Prize in 2023
◆Announcement of the Awardees of the 2023 Neuroscience Research (NSR) Paper Awards.
◆Awardee of the 2023 Joseph Altman Award in Developmental Neuroscience
◆The Winners of the Brain Prize 2023
◆The list of councilors is now available.
◆THE BRAIN PRIZE 2023 WINNERS ANNOUNCED ON 23RD MARCH(CET)/ 24TH MARCH(JST)
◆Message from the president
◆The Brain Prize Webinar – Circuits for Movement
◆Awardee of 2022 Joseph Altman Award in Developmental Neuroscience, Denis Jabaudon
◆Announcement of the 2022 JNS-CNS Travel Award Program Awardees
◆Winners for the 22nd Japan Neuroscience Society Young Investigator Award Selected!
◆Announcement of the 24th Recipients of the Tokizane Award in 2022
◆Announcement of the Awardee of the 2022 Joseph Altman Award in Developmental Neuroscience
◆The Winners of the Brain Prize 2022
◆Announcement of the Awardees of the 2022 Neuroscience Research (NSR) Paper Awards.
◆THE BRAIN PRIZE 2022 WINNERS ANNOUNCED ON 3RD MARCH(CET)/ 4TH MARCH(JST)
◆Announcement of FENS Forum 2022 Travel Awardees
◆Call for SfN2022 Session Proposals
◆Virtual SfN 2021: Global Neuroscience Social
◆Announcement of 2021 JNS-SfN Exchange Travel Awardees
◆Awardees of the 2021 Japan Neuroscience Society Young Investigator Award
◆The Japan Neuroscience Society' s official Journal Neuroscience Research presents NSR Paper Awards
◆Awardee of 2021 Joseph Altman Award in Developmental Neuroscience, Sergiu P. Pasca
◆Announcement of the 23rd Recipient of the Tokizane Prize in 2021
◆Announcement of the Awardee of the 2021 Joseph Altman Award in Developmental Neuroscience
◆Our website has become smartphone friendly.
◆Message from the President
◆JNS is proud to endorse the ALBA Declaration on Equity and Inclusion.
◆Call for Papers for a Special Issue of Brain Structure and Function
◆Awardee of 2020 Joseph Altman Award in Developmental Neuroscience, Haruki Takeuchi
◆Announcement of the Awardee of the 2020 Joseph Altman Award in Developmental Neuroscience
◆JNS join the fight against COVID-19 pandemic
◆Announcement of the 22nd Recipient of the Tokizane Prize in 2020
◆Announcement of 2019 JNS-SfN Exchange Travel Awardees
◆Special Rates Offer for FENS Forum 2020 registration
◆Election of Panel Directors
◆The JNS Secretariat Desk at NEURO2019
◆Announcement of the 2019 Japan Neuroscience Society Young Investigator Awardees
◆Yasunori Hayashi
The 21st Recipient of the Tokizane Prize in 2019
◆Official notice about the election of Panel Directors
◆Altman Award in Developmental Neuroscience
◆Altman Award in Developmental Neuroscience
◆Announcement of the Awardee of the 2019 Joseph Altman Award in Developmental Neuroscience
◆Announcement of the 21st Recipient of the Tokizane Prize in 2019
◆Cast your vote for the new JNS official character!!
◆Special Rates Offer for 2019 ANS Conference,
Australasian Neuroscience Society - 39th Annual Scientific
Meeting registration
◆Altman Award in Developmental Neuroscience
◆Announcement of the Awardee of the 2018 Joseph Altman Award in Developmental Neuroscience
◆Participating report of the 12th biennial Conference of CNS : Kai Li
◆Participating report of the 12th biennial Conference of CNS : Kelvin Hui
◆Itaru Imayoshi the First Awardee of the Altman Award in Developmental Neuroscience
◆Announcement of the First Awardee of the Altman Award in Developmental Neuroscience
◆2020 Student Member Re-Registration and Junior
Member Registration
◆Announcement of the 2025 JNS-CNS Travel Award Program Awardees
◆[Publication] Neuroscience News FY2025 No.1
◆Awardee of 2025 Joseph Altman Award in Developmental Neuroscience, Tomohisa Toda
◆ Announcement of the 27th Recipient of the Tokizane Award in 2025
◆Announcement of the Awardees of the 2025 Neuroscience Research (NSR) Paper Awards.
◆Awardee of the 2025 Joseph Altman Award in Developmental Neuroscience
◆Promotional video for Neuroscience Research is now available on our website.
◆The Winners of the Brain Prize 2025
◆THE BRAIN PRIZE 2025 WINNERS ANNOUNCED ON MARCH 5TH (CET)
◆[Publication] Neuroscience News FY 2024 No.4
◆Participating Report of the 17th Annual Meeting of Chinese Neuroscience Society (CNS 2024)
◆“Meet the Editors” Session (July 24, 25 and 26, 2024) will be held at NEURO2024
◆Neuroscience Research Special Issue(Vol.197) is available for free now
◆Neuroscience Research is now Full Gold Open Access journal
◆Message from the President
◆Atsushi Nambu, The 26th Recipient of the Tokizane Award in 2024
◆Awardee of 2024 Joseph Altman Award in Developmental Neuroscience, Shawn Sorrells
◆Announcement of the 26th Recipients of the Tokizane Prize in 2024
◆Announcement of FENS Forum 2024 Travel Award Awardees
◆Announcement of the 2024 JNS-CNS Travel Award Program Awardees
◆Awardee of the 2024 Joseph Altman Award in Developmental Neuroscience
◆The Winners of the Brain Prize 2024
◆THE BRAIN PRIZE 2024 WINNERS ANNOUNCED ON MARCH 5TH (CET)/ 6TH MARCH(JST)
◆FENS Summer School 2023 Report (Owusu Mensah Richard Nana Abankwah)
◆Call for applications: Chief Editors of Neuroscience
◆Message from the President 'Our Neuroscience Society'
◆Awardee of 2023 Joseph Altman Award in Developmental Neuroscience, Tomasz Nowakowski
◆Announcement of the 24th Recipients of the Tokizane Prize in 2023
◆Announcement of the Awardees of the 2023 Neuroscience Research (NSR) Paper Awards.
◆Awardee of the 2023 Joseph Altman Award in Developmental Neuroscience
◆The Winners of the Brain Prize 2023
◆The list of councilors is now available.
◆THE BRAIN PRIZE 2023 WINNERS ANNOUNCED ON 23RD MARCH(CET)/ 24TH MARCH(JST)
◆Message from the president
◆The Brain Prize Webinar – Circuits for Movement
◆Awardee of 2022 Joseph Altman Award in Developmental Neuroscience, Denis Jabaudon
◆Announcement of the 2022 JNS-CNS Travel Award Program Awardees
◆Winners for the 22nd Japan Neuroscience Society Young Investigator Award Selected!
◆Announcement of the 24th Recipients of the Tokizane Award in 2022
◆Announcement of the Awardee of the 2022 Joseph Altman Award in Developmental Neuroscience
◆The Winners of the Brain Prize 2022
◆Announcement of the Awardees of the 2022 Neuroscience Research (NSR) Paper Awards.
◆THE BRAIN PRIZE 2022 WINNERS ANNOUNCED ON 3RD MARCH(CET)/ 4TH MARCH(JST)
◆Announcement of FENS Forum 2022 Travel Awardees
◆Call for SfN2022 Session Proposals
◆Virtual SfN 2021: Global Neuroscience Social
◆Announcement of 2021 JNS-SfN Exchange Travel Awardees
◆Awardees of the 2021 Japan Neuroscience Society Young Investigator Award
◆The Japan Neuroscience Society' s official Journal Neuroscience Research presents NSR Paper Awards
◆Awardee of 2021 Joseph Altman Award in Developmental Neuroscience, Sergiu P. Pasca
◆Announcement of the 23rd Recipient of the Tokizane Prize in 2021
◆Announcement of the Awardee of the 2021 Joseph Altman Award in Developmental Neuroscience
◆Our website has become smartphone friendly.
◆Message from the President
◆JNS is proud to endorse the ALBA Declaration on Equity and Inclusion.
◆Call for Papers for a Special Issue of Brain Structure and Function
◆Awardee of 2020 Joseph Altman Award in Developmental Neuroscience, Haruki Takeuchi
◆Announcement of the Awardee of the 2020 Joseph Altman Award in Developmental Neuroscience
◆JNS join the fight against COVID-19 pandemic
◆Announcement of the 22nd Recipient of the Tokizane Prize in 2020
◆Announcement of 2019 JNS-SfN Exchange Travel Awardees
◆Special Rates Offer for FENS Forum 2020 registration
◆Election of Panel Directors
◆The JNS Secretariat Desk at NEURO2019
◆Announcement of the 2019 Japan Neuroscience Society Young Investigator Awardees
◆Yasunori Hayashi
The 21st Recipient of the Tokizane Prize in 2019
◆Official notice about the election of Panel Directors
◆Altman Award in Developmental Neuroscience
◆Altman Award in Developmental Neuroscience
◆Announcement of the Awardee of the 2019 Joseph Altman Award in Developmental Neuroscience
◆Announcement of the 21st Recipient of the Tokizane Prize in 2019
◆Cast your vote for the new JNS official character!!
◆Special Rates Offer for 2019 ANS Conference,
Australasian Neuroscience Society - 39th Annual Scientific
Meeting registration
◆Altman Award in Developmental Neuroscience
◆Announcement of the Awardee of the 2018 Joseph Altman Award in Developmental Neuroscience
◆Participating report of the 12th biennial Conference of CNS : Kai Li
◆Participating report of the 12th biennial Conference of CNS : Kelvin Hui
◆Itaru Imayoshi the First Awardee of the Altman Award in Developmental Neuroscience
◆Announcement of the First Awardee of the Altman Award in Developmental Neuroscience
◆2020 Student Member Re-Registration and Junior
Member Registration
Atsushi Nambu, The 26th Recipient of the Tokizane Award in 2024
Movement control functions of the basal ganglia, pathophysiology of their disorders, and therapeutic applications
Atsushi Nambu
Division of Behavioral Development, National Institute for Physiological Sciences
Division of Behavioral Development, National Institute for Physiological Sciences
It is my great pleasure and honor to get an award named after Prof. Toshihiko Tokizane, who was a pioneer in the field of neurophysiology in Japan and the author of “Nou no hanashi (the tale of the brain)”, which I read in my youth.
When I was an undergraduate student in the late 1970’s, it was already known that the basal ganglia played important roles in controlling voluntary movements, as their dysfunctions caused movement disorders, such as Parkinson’s disease. However, their functions and dysfunctions were not well understood. Prof. Kazuo Sasaki, my mentor in the graduate university, suggested me to pursue research on the basal ganglia, because at that time, the basal ganglia were left uninvestigated, unlike the cerebral cortex and cerebellum. Since then, I started research in the field of the basal ganglia.
Following my research training in acute experiments under anesthesia, I utilized electrical stimulation in chronic awake experiments using behaving monkeys. When investigating neuronal activity of the output station of the basal ganglia, the internal segment of the globus pallidus (GPi), I first identified cortical inputs in GPi neurons by response to electrical stimulation in the cerebral cortex, and then recorded neuronal activity during motor tasks. In these recordings, I noticed that GPi neurons showed a triphasic response composed of early excitation, inhibition, and following late excitation in response to cortical stimulation. I assumed that the early excitation could be mediated by the subthalamic nucleus (STN).
Then, after two years of research in slice experiments in the basal ganglia at New York University, I started systematic examination of cortical inputs to the STN by using electrophysiological and anatomical methods with Dr. Masahiko Takada and others in monkeys. We found that the primary motor cortex and supplementary motor area directly projected to the STN in keeping with the somatotopic organization, and named this pathway as the “hyperdirect pathway”, which was distinct from trans-striatal pathways, namely the direct and indirect pathways. Moreover, pharmacological blockade of each pathway revealed that early excitation, inhibition, and late excitation in the GPi induced by cortical stimulation were mediated by the hyperdirect, direct, and indirect pathways, respectively.
We supposed that these pathways could work when executing voluntary movements as well, and proposed the "Dynamic activity model" of the basal ganglia functions: First, signals through the hyperdirect pathway reset on-going cortical activity; second, those through the direct pathway release necessary movements by disinhibiting thalamus; and finally, those through the indirect pathway stop movements.
On the other hand, malfunctions of the basal ganglia cause movement disorders, such as Parkinson’s disease and dystonia. We considered that their pathophysiology could be explained by changes in triphasic response patterns in the GPi. Until then, pathophysiology of movement disorders had been explained by firing rate or firing pattern changes of the basal ganglia, but firing rate changes were not obvious, and firing pattern changes did not properly explain their pathophysiology.
We recorded neuronal activity of parkinsonian monkeys as a starter. Treatment of monkeys with dopaminergic neurotoxin, MPTP, induced motor signs similar to those in human patients. We found that cortically induced inhibition in the GPi was reduced in parkinsonian monkeys (see Figure). This could be interpreted that in the normal state, cortically induced inhibition in the GPi releases movements by disinhibiting thalamus, while in the parkinsonian state, reduced inhibition in the GPi cannot release movements, resulting in akinesia/bradykinesia.
In contrast to Parkinson's disease, there are other movement disorders exhibiting involuntary movements, such as dystonia. We recorded neuronal activity from a mouse model of dystonia, which over-expresses human abnormal proteins, and we found that cortically induced inhibition was enlarged, and late excitation was reduced in the GPi (see Figure). This could be interpreted that enlarged inhibition releases excessive movements, and reduced late excitation is not enough to stop movements, resulting in involuntary movements.
Moreover, making a small lesion or applying continuous electrical stimulation in the basal ganglia ameliorate motor symptoms of advanced Parkinson’s disease. Likewise, pharmacological blockade of STN activity in parkinsonian monkeys recovered cortically induced inhibition in the GPi and improved akinesia/bradykinesia (see Figure). This could be interpreted that recovered inhibition in the GPi can release movements again.
Thus, the "Dynamic activity model" can explain not only the pathophysiology of movement disorders, but also the therapeutic mechanism of stereotactic neurosurgery. If we can find methods to normalize altered triphasic response patterns in movement disorders, that could lead to new therapeutic tools, and we will pursue the development of new tools in the future.
Lastly, I would like to send my deep appreciation to my mentors, colleagues, collaborators, and graduate students who have worked together with me during my research career.
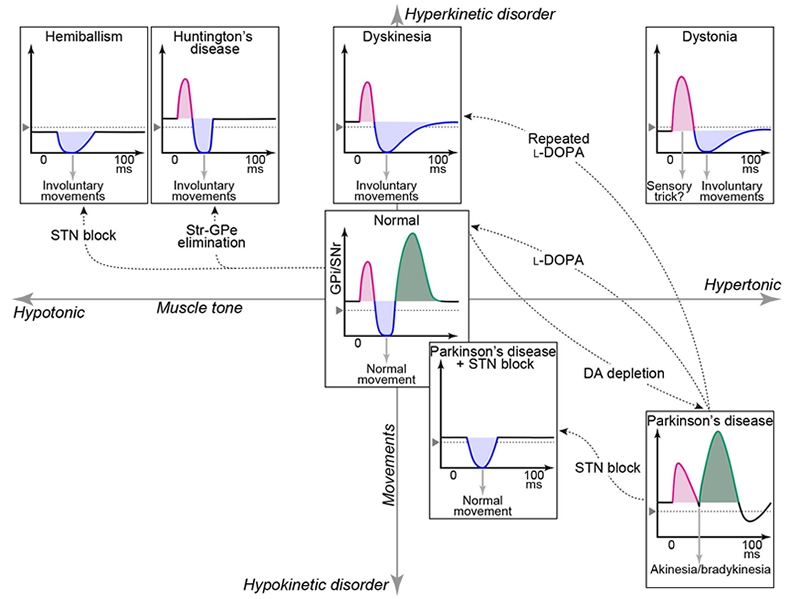
"Dynamic activity model" of movement disorders
Cortically induced response patterns in the output nuclei of the basal ganglia, the internal segment of the globus pallidus and substantia nigra pars reticulata (GPi/SNr), in various movement disorder models are plotted along hyperkinetic-hypokinetic (ordinate) and hypertonic-hypotonic (abscissa) axes. Alterations of cortically induced dynamic activity in the GPi/SNr could explain the pathophysiology of movement disorders in a unified manner. This model could also explain the therapeutic mechanism of stereotactic surgery targeting the subthalamic nucleus (STN) in Parkinson’s disease.
Related References
Nambu A et al. (1996) J Neurosci 16: 2671
Nambu A et al. (2000) J Neurophysiol 84:289
Nambu A et al (2002) Neurosci Res 2002;43:111
Chiken S et al. (2008) J Neurosci 28:13967
Nishibayashi H et al. (2011) Mov Disord 26:469
Sano H et al (2013) J Neurosci 33:7583
Chiken S et al. (2013) J Neurosci 33:2268
Chiken S et al (2015) Cereb Cortex 25:4885
Koketsu D et al. (2021) J Neurosci 41:5502
Chiken S et al (2021) Cereb Cortex 31:5363
Dwi Wahyu I et al. (2021) J Neurosci 41:2668
Darbin O et al. (2022) Sci Rep 12:6493
Hasegawa T et al. (2022) Nat Commun 13:2233.
Nambu A et al (2023) Mov Disord 38: 2145
Nambu A et al. (2000) J Neurophysiol 84:289
Nambu A et al (2002) Neurosci Res 2002;43:111
Chiken S et al. (2008) J Neurosci 28:13967
Nishibayashi H et al. (2011) Mov Disord 26:469
Sano H et al (2013) J Neurosci 33:7583
Chiken S et al. (2013) J Neurosci 33:2268
Chiken S et al (2015) Cereb Cortex 25:4885
Koketsu D et al. (2021) J Neurosci 41:5502
Chiken S et al (2021) Cereb Cortex 31:5363
Dwi Wahyu I et al. (2021) J Neurosci 41:2668
Darbin O et al. (2022) Sci Rep 12:6493
Hasegawa T et al. (2022) Nat Commun 13:2233.
Nambu A et al (2023) Mov Disord 38: 2145

Atsushi Nambu
Division of Behavioral Development, National Institute for Physiological Sciences
Division of Behavioral Development, National Institute for Physiological Sciences
Short CV
| 1982 | Graduate School, Faculty of Medicine, Kyoto University |
| 1985 | Instructor, Faculty of Medicine, Kyoto University |
| 1989 | Postdoctoral Fellow, New York University Medical Center |
| 1991 | Associate Professor, National Institute for Physiological Sciences, |
| 1995 | Staff Scientist, Director, Tokyo Metropolitan Institute for Neuroscience |
| 2002 | Professor, National Institute for Physiological Sciences, The Graduate University for Advanced Studies (SOKENDAI) |
| 2023 | Professor Emeritus, Researcher, National Institute for Physiological Sciences |



HOME | The Japan Neuroscience Society
- About Us
- About Us
- Guidelines
- Directors and Committee Members of the Japan Neuroscience Society
- Councilors
- Honorary Members
- Honorary Members : Dr. Masao Ito
- Supporting Members
- Message from the President
- Laboratory list
- Public Outreach
- Brain Bee
- Official Mascot
- News from JNS
- Annual Meetings
- Annual Meeting List
- The JNS Meeting Planner List
- The 48th Annual Meeting of the Japan Neuroscience Society
- NEURO2024
- The 46th annual meeting of the Japan Neuroscience Society
- NEURO2022
- The 44th Annual Meeting of the Japan Neuroscience Society
- The 43rd Annual Meeting of the Japan Neuroscience Society
- NEURO2019
- Neuroscience Research
- Neuroscience Research
- Articles in Press
- Latest Issue
- Back Issues
- Submit Your Paper
- About the Journal
- Online Subscription
(For Members) - Announcements
- Careers & Training & Awards & Funding
- Faculty and Other positions
- Graduate Student Position/Briefing Session
- Grants/Fellowship/TA Programs
- Call for Applicants
- Awards
- Travel Awards
- Academic Events
- Academic Events
- Advocacy
- Guidelines for ethics-related problems with “non-invasive research on human brain function”
- Guidelines for Animal Experimentation in Neuroscience
- Gender Equality, Inclusion and Diversity
- Member Only
- Online Subscription
(For Members) - The JNS Meeting Planner List [Password Required]
- Log in
- Neuroscience News (Past Issues)
- How to join
- About Membership
- Admission procedure
- Membership Categories